
ฮอร์โมนพืช

ฮอร์โมนพืชเป็นสารเคมีภายในพืชซึ่งเกี่ยวข้องกับการเจริญของพืชไม่เพียงแต่การเจริญของพืชทั้งต้นเท่านั้น หากแต่ยังเกี่ยวข้องกับการเจริญของพืชแต่ละส่วนด้วย ในปัจจุบันทราบกันดีแล้วว่าฮอร์โมนพืชมีทั้งชนิดที่กระตุ้นการเจริญเติบโต และระงับการเจริญเติบโต ฮอร์โมนพืชที่พบในปัจจุบันคือออกซิน (Auxin) จิบเบอเรลลิน (Gibberellins) ไซโตไคนิน(Cytokinins) กรดแอบซิสิค (Abscisic Acid) หรือ ABA และ เอทธิลีน (Ethylene) ซึ่งมีสภาพเป็นก๊าซ
ฮอร์โมนพืช (Phytohormone) คือ สารเคมีที่พืชสร้างขึ้นในปริมาณเพียงเล็กน้อย และ มีผลต่อขบวนการ หรือ ควบคุมการเจริญในพืช (Plant Development)
ฮอร์โมนพืชสามารถเคลื่อนย้ายภายในต้นพืชได้และมีผลกระทบต่อการเจริญเติบโต การเปลี่ยนแปลงทางคุณภาพและการพัฒนาของเนื้อเยื่อ และอวัยวะของพืชซึ่งได้รับฮอร์โมนนั้น ๆ คำว่า ฮอร์โมน นั้นเริ่มใช้โดยนักสรีรวิทยาของสัตว์ ซึ่งต่อมานักสรีรวิทยาของพืชได้นำมาใช้กับสารประกอบอินทรีย์ ซึ่งสามารถมีผลกระทบในปริมาณที่น้อยมาก โดยพืชจะสังเคราะห์ที่ส่วนหนึ่งแล้วเคลื่อนย้ายไปยังอีกส่วนหนึ่ง และมีผลต่อกระบวนการทางสรีรวิทยาที่ค่อนข้างเฉพาะเจาะจง ดังนั้นในการศึกษาทางด้านฮอร์โมนจึงมักศึกษาในแง่ของแหล่งและกระบวนการสังเคราะห์ การเคลื่อนที่และเคลื่อนย้าย และปฏิกิริยาของฮอร์โมนที่มีต่อพืช
สารควบคุมการเจริญเติบโต (Plant Growth Regulator) เป็นสารเคมีที่สำคัญในการเกษตร เป็นสารอินทรีย์ซึ่งมนุษย์สังเคราะห์ขึ้นมาได้ ซึ่งบางชนิดมีคุณสมบัติเหมือนฮอร์โมนพืช มนุษย์รู้จักการใช้สารควบคุมการเจริญเติบโตมานานแล้ว เช่น กระตุ้นให้มะม่วงหรือสับปะรดออกดอกโดยการจุดไฟข้างสวน เพื่อให้เกิดควันซึ่งมีเอทธิลีนปนอยู่ สามารถกระตุ้นให้เกิดการออกดอกได้ ถึงแม้ว่าในขณะนั้นจะยังไม่ทราบสาเหตุที่แท้จริงก็ตาม
การแสดงออกถึงลักษณะต่าง ๆ ของพืชจะเกิดจากพันธุกรรมและสภาพแวดล้อม ซึ่งรวมกับฮอร์โมนด้วย เช่น การปลูกผักกาดขาวปลีบางพันธุ์ในฤดูร้อน ผักกาดขาวปลีจะไม่เข้าหัว ฮอร์โมนบางชนิดสามารถกระตุ้นให้ผักกาดขาวปลีเข้าหัวได้ ซึ่งในกรณีนี้ฮอร์โมนจะทดแทนสภาพแวดล้อมที่เหมาะสมได้
ในประเทศไทยการใช้ฮอร์โมนพืชมีวัตถุประสงค์ในทางการเกษตรเพื่อให้มีผลผลิต เพื่อเพิ่มผลผลิตและคุณภาพ และเพื่อความสะดวกในการจัดการฟาร์ม
ปัจจุบัน จะแบ่งฮอร์โมนพืชออกเป็น
- ออกซิน (Auxins) เป็นสารสังเคราะห์ที่ใช้เร่งราก เปลี่ยนเพศดอก ป้องกันผลร่วง หรือจำกัดวัชพืช ได้แก่ IAA NAA (PlanofixR) IBA (SeradixR)2, 4-D และ 4-CPA
- จิบเบอเรลลิน (Gibberellins) สกัดจากเชื้อราบางชนิด ควบคุมการยืดตัวของเซลล์ การติดผล การเกิดดอก เร่งการเจริญเติบโต ทำลายการพักตัวของเมล็ด ได้แก่ GA ที่ใช้กันมากในปัจจุบันคือ GA3
- ไซโตไคนิน (Cytokinins) ควบคุมการแบ่งเซลล์ การเจริญเติบโตด้านกิ่งใบ การแตกแขนง ชะลอการแก่ ได้แก่ kinetin (SEA-BORNR), BAP
- เอทิลีนและสารปลดปล่อยเอทิลีน (Etylene and Etylene releasing compounds) สารเอทืลีนเป็นก๊าซ ปัจจุบันพัฒนาเป็นของแข็ง ใช้ในการาเร่งดอกสับปะรด เร่งการสุกของผลไม้ เร่งการไหลของน้ำยาง ทำลายการพักตัว ได้แก่สาร ethephon (EthrelR) และ acetylene (เกิดจากถ่านก๊าซ calcium carbide กับน้ำ)
- ชะลอการเจริญเติบโต (Plant Growth Retadants) ไม่พบตามธรรมชาติในพืช ทำงารตรงกันข้ามกับ GA ใช้ชะลอการเจริญเติบโต เช่น ลดความสูงของต้นทำให้ปล้องสั้นลง เพิ่มผลผลิตผัก ป้องกันการหกล้มของธัญพืช ได้แก่ chlomequat (CycocelR), daminozide (AlarR85), mepiquat chlorise (PixR), paxlobutrazol (CultarR)
- สารยับยั้งการเจริญเติบโต (Plant Growth Inhibitors) ใช้ควบคุมไม่ให้พืชโตมากเกินไป นอกจากนี้ hydrazide (OZO-MHR) chlofurenol หรือ morphactin, dikegulac-sodium ใช้เพิ่มการแตกกาข้าง ป้องกันการงอกของมันฝรั่ง หอมหัวใหญ่ ป้องกันการเกิดหน่อของยาสูบ และสารอื่นๆ
- Foleisteine (ErgostimR) กระตุ้นการงอกของเมล็ด เพิ่มการติดผลและเพิ่มผลผลิตของพืชหลายชนิด เช่น ข้าว ข้าวโพด ข้าวสาลี องุ่น ส้ม สตอเบอรี่
- Glyphosine (PolarisR) เร่งการแก่ของอ้อย เพิ่มน้ำตาล
- Glyphosate (RoundupR) เร่งการแก่ของอ้อย เพิ่มน้ำตาลเป็นยากำจัดวัชพืชประเภทยาดูดซึม
- Paraqyet (GranoxomeR) ทำลายส่วนสีเขียว ทำให้ใบร่วง
- Sodium mono-nitro-quaiacol (AtonikR) ใช้เร่งการเจริญเติบโต เพิ่มการติดผล ป้องกันใบร่วง
ออกซิน (Auxin)
ความรู้พื้นฐานเกี่ยวกับออกซินนั้นเกิดขึ้นจากงานของ Charles Darwin ซึ่งศึกษาเรื่อง Phototropism ซึ่งพืชจะโค้งงอเข้าหาแสง Darwin ทดลองกับต้นกล้าของ Phalaris canariensis และพบว่าโคลีออพไทล์ของพืชชนิดนี้จะตอบสนองต่อการได้รับแสงเพียงด้านเดียวทำให้เกิดการโค้งเข้าหาแสง Darwin สรุปว่าเมื่อต้นกล้าได้รับแสงจะทำให้มี “อิทธิพล” (Influence) บางอย่างส่งผ่านจากส่วนยอดมายังส่วนล่างของโคลีออพไทล์ ทำให้เกิดการโค้งงอเข้าหาแสง ซึ่งนักวิทยาศาสตร์รุ่นต่อมาได้ศึกษาถึง “อิทธิพล” ดังกล่าว
ต่อมา Boysen-Jensen และ Paal ได้ศึกษาและแสดงให้เห็นว่า “อิทธิพล” ดังกล่าวนี้มีสภาพเป็นสารเคมี ซึ่งในสภาพที่โคลีออพไทล์ได้รับแสงเท่ากันทั้งสองด้าน สารเคมีนี้จะเคลื่อนที่ลงสู่ส่วนล่างของโคลีออพไทล์ ในอัตราเท่ากันทุกด้านและทำหน้าที่เป็นสารกระตุ้นการเจริญเติบโต
ในปี ค.ศ. 1926 Went ได้ทำงานทดลองและสามารถแยกสารชนิดนี้ออกจากโคลีออพไทล์ได้ โดยตัดส่วนยอดของโคลีออพไทล์ของข้าวโอ๊ตแล้ววางลงบนวุ้นจะทำให้สารเคมีที่กระตุ้นการเจริญเติบโตไหลลงสู่วุ้น เมื่อนำวุ้นไปวางลงที่ด้านหนึ่งของโคลีออพไทล์ที่ไม่มียอดด้านใดด้านหนึ่งจะทำให้โคลีออพไทล์ดังกล่าวโค้งได้ เขาสรุปว่าสารเคมีได้ซึมลงสู่วุ้นแล้วซึมจากวุ้นลงสู่ส่วนของโคลีออพไทล์ วิธีการดังกล่าวนอกจากเป็นวิธีการแรกที่แยกสารเคมีชนิดนี้ได้แล้ว ยังเป็นวิธีการวัดปริมาณของฮอร์โมนได้ด้วย เป็นวิธีที่เรียกว่า Bioassay
สารเคมีดังกล่าวได้รับการตั้งชื่อว่า ออกซิน ซึ่งในปัจจุบันพบในพืชชั้นสูงทั่วๆ ไป และมีความสำคัญต่อการเจริญเติบโตของพืช สังเคราะห์ได้จากส่วนเนื้อเยื่อเจริญของลำต้น ปลายราก ใบอ่อน ดอกและผล และพบมากที่บริเวณเนื้อเยื่อเจริญ โคลีออพไทด์และคัพภะ รวมทั้งใบที่กำลังเจริญด้วย

กลไกการทำงานของออกซิน
โดยทั่วไปฮอร์โมนจะสามารถก่อให้เกิดผลต่อการเจริญเติบโตได้ในปริมาณที่ต่ำมาก จึงสรุปกันว่าการทำงานของฮอร์โมนต้องเกี่ยวข้องกับการขยายสัญญาณของฮอร์โมน (Large Amplification) แล้วฮอร์โมนสามารถทำให้เกิดการเปลี่ยนแปลงของโมเลกุลจำนวนมากขึ้นได้ โดยทั่วไปฮอร์โมนจะมีผลต่อการเจริญเติบโตโดยผ่านมาทางการควบคุมการสังเคราะห์โปรตีนหรือกรดนิวคลีอิคควบคุม “pace-setter” ของเอนไซม์และควบคุมการยอมให้สารเข้าออกจากเซลล์ของเยื่อหุ้มเซลล์
กลไกในการทำงานของออกซินในระยะที่ผ่านมาจะมีแนวความคิดเป็นสองอย่าง คือ แนวคิดที่เกี่ยวข้องกับผนังเซลล์เป็นส่วนที่รับผลกระทบของออกซินและขยายตัว ส่วนอีกแนวคิดหนึ่งมุ่งไปที่ผลของออกซินต่อเมตาบอลิสม์ของกรดนิวคลีอิค ในปัจจุบันได้นำสองแนวคิดมาวิเคราะห์ ร่วมกันเพื่อศึกษากลไกในการทำงานของออกซิน และยังศึกษาผลของออกซินต่อเยื่อหุ้มเซลล์ด้วย
การขยายตัวของเซลล์จะสัมพันธ์กับการเปลี่ยนแปลงปริมาณและกิจกรรมของเอนไซม์ โดยที่ออกซินจะมีบทบาทต่อ กระบวนการเมตาบอลิสม์ของกรดนิวคลีอิค โดยการศึกษาจากการเพาะเลี้ยงเนื้อเยื่อที่เป็นไส้ของต้นยาสูบ (Tobacco Pith) ซึ่งจะเจริญไปเป็นกลุ่มเนื้อเยื่อ (Callus) นั้นพบว่ามีปริมาณของ RNA เพิ่มมากขึ้น ทั้งนี้เพราะออกซินจะกระตุ้นให้มีการสังเคราะห์ RNA มากขึ้น แล้วส่งผลไปถึงการเจริญของกลุ่มเนื้อเยื่อ ถ้าหากใช้สารระงับการสังเคราะห์โปรตีนหรือ RNA ความสามารถในการกระตุ้นการเจริญเติบโตของออกซินจะหายไป
การทดลองอีกเรื่องที่ชี้ให้เห็นว่าออกซินกระตุ้นให้มีการสร้าง RNA คือ การใช้นิวเคลียส หรือโครมาตินเลี้ยงไว้ในสารที่เป็นสารเริ่มต้นของ RNA เช่น ATP CTP GTP และ UTP ซึ่งสารเริ่มต้นเหล่านี้จะมีสารกัมมันตรังสีปรากฏอยู่ด้วย RNA ที่เกิดขึ้นมาใหม่จะมีสารกัมมันตรังสีด้วย ซึ่งการที่จะเกิด RNA ใหม่ขึ้นได้นี้เซลล์จะต้องได้รับออกซินก่อนที่นิวเคลียสหรือโครมาตินจะถูกแยกออกจากเซลล์เท่านั้น ซึ่งแสดงให้เห็นว่าออกซินไปกระตุ้นการสังเคราะห์ RNA
ดังนั้นจึงเป็นที่ชัดเจนว่าออกซินมีผลต่อระดับเอนไซม์ โดยผ่านทางการสังเคราะห์ RNA นอกจากนั้นออกซินยังมีผลกระทบต่อกิจกรรมของเอนไซม์โดยตรง เช่น การกระตุ้นให้เอนไซม์เกิดกิจกรรมหรือเปลี่ยนรูปมาอยู่ในรูปที่มีกิจกรรมได้ แต่ไม่ว่าออกซินจะมีผลกระทบต่อเอนไซม์แบบใดก็ตาม นักวิทยาศาสตร์ได้มุ่งความสนใจไปสู่เอนไซม์ที่สัมพันธ์กับกระบวนการขยายตัวของเซลล์ เซลล์พืชจะมีผนังเซลล์อยู่ข้างนอกสุด ดังนั้นการเจริญของเซลล์จะเกิดขึ้นได้เมื่อคุณสมบัติของผนังเซลล์เปลี่ยนไปในทางที่ก่อให้เกิดการขยายตัวของโปรโตพลาสต์ จากความจริงดังกล่าวการศึกษาทางด้านนี้จึงมุ่งไปสู่ผลกระทบของออกซินต่อคุณสมบัติของผนังเซลล์
เซลล์พืชทุกชนิดที่ผ่านขั้นตอนของเนื้อเยื่อเจริญมาแล้วจะผ่านขั้นตอนการ เจริญเติบโต 2 ขั้น คือ การแบ่งเซลล์และการขยายตัวแวคคิวโอขึ้นภายในเซลล์ (Vacuolation) ในการศึกษาการเจริญเติบโตของโคลีออพไทล์ของข้าวโอ๊ต พบว่าการแบ่งเซลล์จะหยุดเมื่อมีความยาว 10 มิลลิเมตร การเจริญเติบโตที่เกิดขึ้นหลังจากนั้นจะเนื่องมาจากการขยายขนาดของเซลล์ ดังนั้นในการศึกษาถึงผลกระทบของออกซินต่อการเจริญเติบโตของพืชจึงเน้นไปที่ผลต่อการขยายตัวของเซลล์ ในระหว่างการขยายขนาดของเซลล์เพราะการขยายตัวของแวคคิวโอ หรืออาจจะเกิดช่องว่างภายในเซลล์ขึ้น ที่ผนังเซลล์จะเกิดการยืดตัวชนิดที่ไม่สามารถหดได้อีก มีการทดลองหลายการทดลองสนับสนุนว่าออกซินเพิ่มการยืดตัวของผนังเซลล์ (Plasticity)
ในระหว่างการขยายตัวของเซลล์นั้น ไม่เพียงแต่ผนังเซลล์ยืดตัวเท่านั้น แต่ยังมีการเพิ่มความหนาของผนังเซลล์เพราะมีสารใหม่ ๆ ไปเกาะด้วย ซึ่งการเจริญดังกล่าวนี้ก็เป็นผลจากการกระตุ้นของออกซิน ซึ่งจะเกิดขึ้นเมื่อการยืดตัวของเซลล์หยุดลงแล้ว
ผนังเซลล์ประกอบด้วยเซลล์ลูโลสไมโครไฟบริลฝังตัวอยู่ในส่วนที่เป็นแมททริกซ์ (Matrix) และโปรตีน ดังนั้นถ้าพิจารณาดูผนังเซลล์จะมีลักษณะเหมือนคอนกรีตเสริมเหล็กโดยเซลลูโลสจะเป็นส่วนของเหล็ก โมเลกุลของเซลลูโลสยึดติดกันด้วยแขนไฮโดรเจน (Hydrogen bond) ในขณะที่ส่วนของแมททริกซ์เกาะกันด้วยแขนโควาเลนท์ (Covalent bond) และในเนื้อเยื่อใบเลี้ยงของพืชใบเลี้ยงคู่ เซลลูโลสเกาะอยู่กับแมททริกซ์โดยแขนไฮโดรเจน ดังนั้นการที่ออกซินจะทำให้เซลล์ยืดตัวนั้นต้องทำลายแขนเหล่านี้เสียก่อน ในปัจจุบันเป็นที่ทราบกันแล้วว่าแม้ว่าการขยายตัวของเซลล์เกี่ยวข้องกับการสังเคราะห์โปรตีนและ RNA รวมทั้งพลังงานจากการหายใจก็ตาม แต่ถ้าให้ออกซินจากภายนอกต่อลำต้นหรือโคลีออพไทล์อัตราการเจริญเติบโตจะเพิ่มขึ้นหลังจากระยะเวลา “lag” เพียง 2-3 นาทีเท่านั้น ซึ่งเป็นไปไม่ได้ที่การเจริญเติบโตถูกเร่งโดยการเปลี่ยนอัตราของการ Transcription และ Translation แต่ดูเหมือนว่าออกซินจะไปมีผลต่อระบบที่ปรากฏอยู่ในพืชแล้ว (Pre-formed System) ดังนั้นจากเหตุผลข้างต้น
การตอบสนองของพืชต่อออกซิน
- การตอบสนองในระดับเซลล์ ออกซินทำให้เกิดการขยายตัวของเซลล์ (Cell enlargement) เช่น ทำให้เกิดการขยายตัวของใบ ทำให้ผลเจริญเติบโต เช่น กรณีของสตรอเบอรี่ ถ้าหากกำจัดแหล่งของออกซิน ซึ่งคือส่วนของเมล็ดที่อยู่ภายนอกของผล (ผลแห้งแบบ Achene) จะทำให้เนื้อเยื่อของผลบริเวณที่ไม่มีเมล็ดรอบนอกไม่เจริญเติบโต ออกซินทำให้เกิดการแบ่งเซลล์ได้ในบางกรณี เช่น กระตุ้นการแบ่งเซลล์ของแคมเบียมและกระตุ้นให้เกิดการเปลี่ยนแปลงทางคุณภาพ เช่น กระตุ้นให้เกิดท่อน้ำและท่ออาหาร กระตุ้นให้เกิดรากจากการปักชำพืช เช่น การใช้ IBA ในการเร่งรากของกิ่งชำ แล้วยังกระตุ้นให้เกิดแคลลัส (Callus) ในการเพาะเลี้ยงเนื้อเยื่อ แต่การตอบสนองในระดับเซลล์ที่เกิดเสมอคือ การขยายตัวของเซลล์
- การตอบสนองของอวัยวะหรือพืชทั้งต้น
2.1 เกี่ยวข้องกับการตอบสนองของพืชต่อแสง (Phototropism) Geotropism ดังได้กล่าวมาแล้ว
2.2 การที่ตายอดข่มไม่ให้ตาข้างเจริญเติบโต (Apical Dominance)
2.3 การติดผล เช่น กรณีของมะเขือเทศ ออกซินในรูปของ 4 CPA จะเร่งให้เกิดผลแบบ Pathenocarpic และในเงาะถ้าใช้ NAA 4.5 เปอร์เซ็นต์ จะเร่งการเจริญของเกสรตัวผู้ทำให้สามารถผสมกับเกสรตัวเมียได้ ในดอกที่ได้รับ NAA เกสรตัวเมียจะไม่เจริญเพราะได้รับ NAA ที่มีความเข้มข้นสูงเกินไป แต่เกสรตัวผู้ยังเจริญได้ ทำให้การติดผลเกิดมากขึ้น
2.4 ป้องกันการร่วงของผลโดยออกซินจะยับยั้งไม่ให้เกิด Abcission layer ขึ้นมา เช่น การใช้ 2,4-D ป้องกันผลส้มไม่ให้ร่วง หรือ NAA สามารถป้องกันการร่วงของผลมะม่วง
2.5 ป้องกันการร่วงของใบ
2.6 ในบางกรณีออกซินสามารถทำให้สัดส่วนของดอกตัวเมีย และตัวผู้เปลี่ยนไปโดยออกซินจะกระตุ้นให้มีดอกตัวเมียมากขึ้น
จิบเบอเรลลิน (Gibberellins)
การค้นพบกลุ่มของฮอร์โมนพืชที่ปัจจุบันเรียกว่าจิบเบอเรลลินนั้น เกิดประมาณ ปี 1920 เมื่อ Kurosawa นักวิทยาศาสตร์ชาวญี่ปุ่น ศึกษาในต้นข้าวที่เป็นโรค Bakanae หรือโรคข้าวตัวผู้ ซึ่งเกิดจากเชื้อรา Gibberella fujikuroi หรือ Fusarium moniliforme ซึ่งทำให้ต้นข้าวมีลักษณะสูงกว่าต้นข้าวปกติ ทำให้ล้มง่าย จากการศึกษาพบว่า ถ้าเลี้ยงเชื้อราในอาหารเลี้ยงเชื้อเหลวแล้วกรองเอาเชื้อราออกไปเหลือแต่อาหารเลี้ยงเชื้อ เมื่อนำไปราดต้นข้าวจะทำให้ต้นข้าวเป็นโรคได้ จึงเป็นที่แน่ชัดว่า เชื้อราชนิดนี้สามารถสร้างสารบางชนิดขึ้นในต้นพืชหรือในอาหารเลี้ยงเชื้อ ซึ่งกระตุ้นให้ต้นข้าวเกิดการสูงผิดปกติได้ ในปี 1939 ได้มีผู้ตั้งชื่อสารนี้ว่าจิบเบอเรลลิน การค้นพบจิบเบอเรลลิน เกิดขึ้นในช่วงเดียวกับที่พบออกซิน การศึกษาส่วนใหญ่จึงเน้นไปทางออกซิน ส่วนการศึกษาจิบเบอเรลลินในช่วงแรกจะเป็นไปในแง่ของโรคพืช ในการศึกษาขั้นแรกค่อนข้างยากเพราะมักจะมีกรดฟิวซาริค (Fusaric Acid) ปะปนอยู่ซึ่งเป็นสารระงับการเจริญเติบโต ความรู้เกี่ยวกับโครงสร้างและส่วนประกอบทางเคมีของจิบเบอเรลลินนั้นได้รับการศึกษาในปี 1954 โดยนักเคมีชาวอังกฤษซึ่งสามารถแยกสารบริสุทธิ์จากอาหารเลี้ยงเชื้อรา Gibberella fujikuroi และเรียกสารนี้ว่ากรดจิบเบอเรลลิค (Gibberellic Acid)
การให้กรดจิบเบอเรลลิคกับพืชที่สมบูรณ์ทั้งต้น จะเร่งให้เกิดการยืดตัวเพิ่มขึ้นของลำต้นและใบอย่างผิดปกติ การตอบสนองจะปรากฏเด่นชัดเมื่อให้กรดนี้กับพืชที่เตี้ยแคระโดยพันธุกรรม เพราะจะกระตุ้นให้พืชเหล่านี้เจริญสูงตามปกติ กรดจิบเบอเรลลิคที่พบในอาหารเลี้ยงเชื้อรานั้นมีโครงสร้างทางเคมี และกิจกรรมทางชีววิทยาเหมือนกับกรดจิบเบอเรลลิคในพืชปกติทุก ๆ ชนิด (พืชปกติหมายถึงพืชที่ไม่เป็นโรค) มีสารประกอบประเภทนี้จำนวนมากที่แยกเป็นสารบริสุทธิ์ได้จากพืชชั้นสูง ในปัจจุบันมีจิบเบอเรลลินซึ่งเป็นชื่อเรียกทั่ว ๆ ไปของสารประกอบประเภทนี้ ประมาณไม่น้อยกว่า 80 ชนิด ชื่อเรียกสารประกอบชนิดนี้จะตั้งชื่อดังนี้ คือ Gibberellins A1 (GA1), A2, A3 เป็นต้น โดยที่กรดจิบเบอเรลลิค คือ GA3
GA ทุกชนิดจะมีโครงสร้างพื้นฐานของโมเลกุลเป็น Gibberellane Carbon Skeleton ซึ่งจะเหมือนกับกรดจิบเบอเรลลิค จะแตกต่างกันตรงจำนวนและตำแหน่งของกลุ่มที่เข้าแทนที่ในวงแหวนและระดับของความอิ่มตัวของวงแหวน A GA ประกอบด้วยคาร์บอนประมาณ 19-20 อะตอม ซึ่งจะรวมกันเป็นวงแหวน 4 หรือ 5 วงและจะต้องมีกลุ่มคาร์บอกซิลอย่างน้อย 1 กลุ่ม โดยใช้ชื่อย่อว่า GA ซึ่ง GA3 เป็นชนิดที่พบมากและได้รับความสนใจศึกษามากกว่าชนิดอื่นๆ GA เป็นฮอร์โมนที่พบในพืชชั้นสูงทุกชนิด นอกจากนั้นยังพบในเฟิร์น สาหร่าย และเชื้อราบางชนิด แต่ไม่พบในแบคทีเรีย
การเคลื่อนย้ายของจิบเบอเรลลินในต้นพืช
จิบเบอเรลลินสามารถเคลื่อนย้ายหรือเคลื่อนที่ในพืชได้ทั้งทางเบสิพีตัล และ อะโครพีตัล และการเคลื่อนที่ไม่มีโพลาริตี้ การเคลื่อนย้ายเกิดขึ้นทั้งในส่วนของท่ออาหารและท่อน้ำ แต่การเคลื่อนที่ของจิบเบอเรลลินจากยอดอ่อนลงมาสู่ส่วนล่างของลำต้นนั้นไม่ได้เกิดในท่อน้ำ ท่ออาหารเพราะส่วนของยอดอ่อนเป็นส่วนที่ดึงอาหารและธาตุอาหารให้เคลื่อนที่ขึ้นไปแบบ อะโครพีตัล ดังนั้นจิบเบอเรลลินจึงไม่ได้เคลื่อนที่ทางท่ออาหาร และยังไม่ทราบวิถีการเคลื่อนที่แน่ชัด
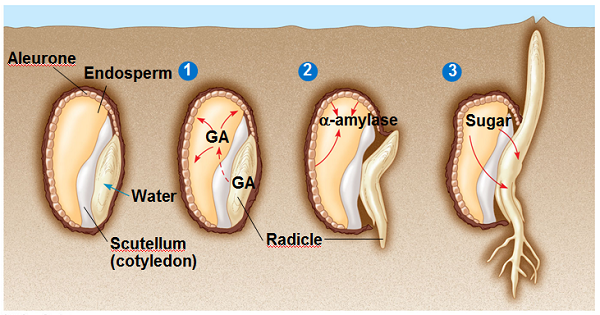
กลไกในการทำงานของจิบเบอเรลลิน การศึกษาด้านกลไกในการทำงานของจิบเบอเรลลินเกิดจากการที่พบว่ามีระดับของกิจกรรมของเอนไซม์หลายชนิดมีผลกระทบจากปริมาณของจิบเบอเรลลิน เอนไซม์ซึ่งมีกิจกรรมเพิ่มขึ้นเมื่อได้รับจิบเบอเรลลิน คือ เอนไซม์ แอลฟาและเบตา-อะมัยเลส (a และ b-amylase) โปรตีเอส (Protease) และไรโบนิวคลีเอส (Ribonuclease) ซึ่งพบในเมล็ดข้าวบาร์เลย์ซึ่งกำลังงอก นอกจากนั้นในพืชบางชนิดยังพบว่ากิจกรรมของไนเตรท รีดักเตส (Nitrate Reductase) และ ไรบูโลสฟอสเฟสคาร์บอกซิเลส (Ribulose Phosphate Carboxylase) มีกิจกรรมเพิ่มขึ้นด้วย ใน ต้นอ้อยนั้น พบว่า ผลของจิบเบอเรลลินจะชะลอการสังเคราะห์อินเวอร์เทส (Invertase) และ เพอรอกซิเดส (Peroxidase) ความสนใจในกลไกการทำงานของจิบเบอเรลลิน จึงเน้นไปที่ การศึกษาว่าจิบเบอเรลลินควบคุมกิจกรรมของเอนไซม์ได้เพราะเป็นผลเนื่องมาจากเปลี่ยนแปลงการสังเคราะห์โปรตีนโดย RNA
การศึกษาตัวอย่างของระดับกิจกรรมของเอนไซม์ซึ่งถูกควบคุมโดยจิบเบอเรลลิน ทำกันมากในเอนไซม์ แอลฟา อะมัยเลส ในเมล็ดข้าวบาร์เลย์ ในเมล็ดข้าวบาร์เลย์ที่แห้งที่ยังไม่ดูดซับน้ำจะไม่มีเอนไซม์แอลฟา อะมัยเลสปรากฏอยู่ เอนไซม์นี้จะปรากฏขึ้นและปลดปล่อยออกมาจากชั้นของอะลีโรนของเมล็ด เป็นการตอบสนองต่อจิบเบอเรลลินซึ่งสังเคราะห์จากต้นอ่อนที่กำลังงอก เนื้อเยื่อชั้นอะลีโรนซึ่งแยกจากเมล็ดที่ไม่งอกจะมีกิจกรรมของแอลฟา อะมัยเลส น้อยมาก แต่ถ้านำเนื้อเยื่อนี้ไปแช่ในจิบเบอเรลลินจะทำให้เกิดการเพิ่มกิจกรรมของแอลฟา อะมัยเลสมากขึ้นโดยเกิดขึ้นหลังจากแช่ไว้นาน 8 ชั่วโมงแล้ว การกระตุ้นให้เกิดการสร้างแอลฟา อะมัยเลสนี้ จะชะงักไปเมื่อใช้สารระงับการสังเคราะห์ RNA และโปรตีน รวมอยู่ในสารละลายจิบเบอเรลลิน ซึ่งจากการทดลองดังกล่าวแสดงว่าจิบเบอเรลลินควบคุมกิจกรรมของแอลฟา อะมัยเลส ผ่านทางการสังเคราะห์ RNA สารชะงักการสังเคราะห์ RNA เช่น แอคติโนมัยซิน-ดี (Actinomycin-D) จะชะงักกระบวนการกระตุ้นการสังเคราะห์ RNA 2-3 ชั่วโมง หลังจากเติมจิบเบอเรลลิน ในขณะที่สารชะงักการสังเคราะห์โปรตีน เช่น ไซโคลเฮคซิไมด์ (Cycloheximide) จะระงับการปรากฏของกิจกรรมของแอลฟา อะมัยเลส หลังจากช่วง “lag” เริ่มต้น
กลไกในการทำงานขั้นแรกของจิบเบอเรลลินนั้นจะเปลี่ยนระบบเยื่อหุ้มเซลล์แล้วจึงจะไปมีผลในการกระตุ้นการสังเคราะห์ RNA และโปรตีน นั่นคือในการกระตุ้นระยะสั้นจะเกี่ยวข้องกับระบบเยื่อหุ้มเซลล์ ในระยะยาวจะเกี่ยวข้องกับการสังเคราะห์ RNA และโปรตีน กลไกที่เกี่ยวข้องกับการเปลี่ยนระบบของเยื่อหุ้ม คือ เพิ่มการสังเคราะห์เยื่อหุ้มทำให้เกิดเอนโดพลาสมิคเรตติคิวลัม มากขึ้น และกระตุ้นการสร้างเวสซิเคิลซึ่งมีเอนไซม์อยู่ภายใน นอกจากนั้นยังกระตุ้นให้มีการปลดปล่อย แอลฟา อะมัยเลส ผ่านเยื่อหุ้มเซลล์ออกมา
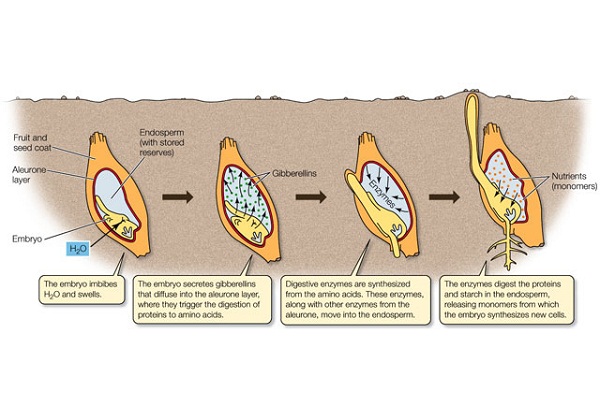
บทบาทของจิบเบอเรลลินที่มีต่อพืช
- กระตุ้นการเจริญเติบโตของพืชทั้งต้น จิบเบอเรลลินมีคุณสมบัติพิเศษ ซึ่งสามารถกระตุ้นการเจริญเติบโตของพืชทั้งต้นได้โดยทำให้เกิดการยืดตัวของเซลล์ ซึ่งผลนี้จะต่างจากออกซินซึ่งสามารถกระตุ้นการเจริญเติบโตของชิ้นส่วนของพืชได้ พืชบางชนิดอาจจะไม่ตอบสนองต่อจิบเบอเรลลิน อาจจะเป็นเพราะว่าในพืชชนิดนั้นมีปริมาณจิบเบอเรลลินเพียงพอแล้ว จิบเบอเรลลินสามารถกระตุ้นการยืดยาวของช่อดอกไม้บางชนิดและทำให้ผลไม้มีรูปร่างยาวออกมา เช่น องุ่น และแอปเปิล กะหล่ำปลีซึ่งเจริญในลักษณะต้นเตี้ยเป็นพุ่ม (Rosette) มีปล้องสั้นมาก เมื่อให้ GA3 กับต้นกะหล่ำปลีดังกล่าวจะทำให้สูงขึ้นถึง 2 เมตรได้ ถั่วพุ่มที่ได้รับ GA จะกลายเป็นถั่วเลื้อยได้ พืชซึ่งมีต้นเตี้ยทางพันธุกรรม เช่น ข้าว ข้าวโพด ถั่ว แตงกวาและแตงโมสามารถแสดงลักษณะปกติได้เมื่อได้รับ GA3 ในข้าวโพดแคระนั้นพบว่าความผิดปกติเกิดจากยีนส์ควบคุม ซึ่งอาจจะเกี่ยวพันกับวิถีในการสังเคราะห์จิบเบอเรลลิน ส่วนข้าวโพดปกติหากได้รับจิบเบอเรลลินจะไม่สามารถสูงขึ้นได้อีก ดังนั้นในกรณีข้าวโพดการแคระเกิดจากมีปริมาณจิบเบอเรลลินในต้นน้อยเกินไป แต่อาการแคระของพืชบางชนิด เช่น Japanese Morning Glory พบว่ามีจิบเบอเรลลินมากพอแล้ว แต่เมื่อได้รับ จิบเบอเรลลินเพิ่มก็จะสูงขึ้นได้ ในกรณีนี้อาจจะเป็นเพราะในต้นมีปริมาณของสารระงับการ เจริญเติบโตอยู่สูง
- กระตุ้นการงอกของเมล็ดที่พักตัวและตาที่พักตัว ตาของพืชหลายชนิดที่เจริญอยู่ในเขตอบอุ่นจะพักตัวในฤดูหนาว เมล็ดของพืชหลายชนิดมีพฤติกรรมเช่นนี้ด้วย ซึ่งการพักตัวจะลดลงจนหมดไป เมื่อได้รับความเย็นเพียงพอ การพักตัวของเมล็ดและตา อันเนื่องมาจากต้องการอุณหภูมิต่ำ วันยาว และต้องการแสงสีแดงจะหมดไปเมื่อได้รับจิบเบอเรลลิน
- การแทงช่อดอก การออกดอกของพืชเกี่ยวข้องกับปัจจัยหลายอย่าง เช่น อายุ และสภาพแวดล้อม จิบเบอเรลลินสามารถแทนความต้องการวันยาวในพืชบางชนิดได้ และยังสามารถทดแทนความต้องการอุณหภูมิต่ำ (Vernalization) ในพืชพวกกะหล่ำปลี และแครอท
- จิบเบอเรลลิน สามารถกระตุ้นการเคลื่อนที่ของอาหารในเซลล์สะสมอาหาร หลังจากที่เมล็ดงอกแล้ว เพราะรากและยอดที่ยังอ่อนตัวเริ่มใช้อาหาร เช่น ไขมัน แป้ง และโปรตีน จากเซลล์สะสมอาหาร จิบเบอเรลลินจะกระตุ้นให้มีการย่อยสลายสารโมเลกุลใหญ่ให้เป็นโมเลกุลเล็ก เช่น ซูโครสและกรดอะมิโน ซึ่งเกี่ยวพันกับการสังเคราะห์เอนไซม์หลายชนิดดังกล่าวข้างต้น
- กระตุ้นให้เกิดผลแบบ Parthenocarpic ในพืชบางชนิด เปลี่ยนรูปร่างของใบพืชบางชนิด เช่น English Ivy และทำให้พืชพัฒนาการเพื่อทนความเย็นได้
- พืชที่มีดอกตัวผู้ และตัวเมียแยกกันไม่ว่าจะต้นเดียวกัน หรือแยกต้นก็ตาม จิบเบอเรลลินสามารถเปลี่ยนเพศของดอกได้ จิบเบอเรลลินมักเร่งให้เกิดดอกตัวผู้ ส่วนออกซิน เอทธิลีน และไซโตไคนิน มักจะเร่งให้เกิดดอกตัวเมีย ในแตงกวาดอกล่าง ๆ มักเป็นดอกตัวผู้ และดอกบนเป็นดอกตัวเมีย การให้สารอีธีฟอนจะเร่งให้เกิดดอกตัวเมียขึ้น
ไซโตไคนิน (Cytokinins)
การค้นพบฮอร์โมนในกลุ่มนี้เริ่มจากการศึกษาการเพาะเลี้ยงเนื้อเยื่อ โดยในปี ค.ศ. 1920 Haberlandt ได้แสดงให้เห็นว่ามีสารชนิดหนึ่งเกิดอยู่ในเนื้อเยื่อพืชและกระตุ้นให้เนื้อเยื่อพาเรนไคมาในหัวมันฝรั่งกลับกลายเป็นเนื้อเยื่อเจริญได้ ซึ่งแสดงว่าสารชนิดนี้สามารถกระตุ้นให้มีการแบ่งเซลล์ ต่อมามีการพบว่าน้ำมะพร้าวและเนื้อเยื่อของหัวแครอทมีคุณสมบัติในการกระตุ้นการแบ่งเซลล์เช่นกัน
นักวิทยาศาสตร์หลายท่าน เช่น Skoog และ Steward ทำการทดลองในสหรัฐอเมริกา โดยศึกษาความต้องการสิ่งที่ใช้ในการเจริญเติบโตของกลุ่มก้อนของเซลล์ (Callus) ซึ่งเป็นเซลล์ที่แบ่งตัวอย่างรวดเร็ว แต่ไม่มีการเปลี่ยนแปลงทางคุณภาพเกิดขึ้นของ pith จากยาสูบและรากของแครอท จากผลการทดลองนี้ทำให้รู้จักไซโตไคนินในระยะปี ค.ศ. 1950 ซึ่งเป็นฮอร์โมนพืชที่จำเป็นต่อการแบ่งเซลล์และการเปลี่ยนแปลงทางคุณภาพของเนื้อเยื่อ ในปัจจุบันพบว่าไซโตไคนิน ยังเกี่ยวข้องกับการเสื่อมสภาพ (Senescence) และการควบคุมการเจริญของตาข้างโดยตายอด (Apical Dominance)
จากการศึกษาของ Skoog โดยเลี้ยงเนื้อเยื่อ pith ของยาสูบ พบว่าการที่เนื้อเยื่อจะเจริญต่อไปได้นั้นจะต้องมีอาหารและฮอร์โมน เช่น ออกซิน โดยถ้าให้ออกซินในอาหารจะมีการเจริญของเนื้อเยื่อนั้นน้อยมาก เซลล์ขนาดใหญ่เกิดขึ้นโดยไม่แบ่งเซลล์ นอกจากนั้นจะไม่เกิดการเปลี่ยนแปลงทางคุณภาพ อย่างไรก็ตามหากเพิ่มพิวรีน เบส (Purine Base) ชนิดอะดีนีน (Adenine) ลงไปในอาหารรวมกับ IAA พบว่า เนื้อเยื่อจะกลายเป็นกลุ่มเซลล์ (Callus) ถ้าใส่อะดีนีนอย่างเดียวรวมกับอาหาร เนื้อเยื่อจะไม่สร้างกลุ่มเซลล์ขึ้นมา ดังนั้นจึงมีปฏิสัมพันธ์ (Interaction) ระหว่าง อะดีนีน และ IAA ซึ่งกระตุ้นให้เกิดการแบ่งเซลล์ขึ้น อะดีนีนเป็นพิวรีนเบส ซึ่งมีสูตรเป็น 6-อะมิโนพิวรีน (6-aminopurine) และปรากฏอยู่ในสภาพธรรมชาติโดยเป็นส่วนประกอบของกรด นิวคลีอิค
ในปี 1955 Miller ได้แยกสารอีกชนิดหนึ่งซึ่งมีคุณสมบัติคล้ายคลึงแต่มีประสิทธิภาพดีกว่าอะดีนีน ซึ่งได้จากการสลายตัวของ DNA ของสเปิร์มจากปลาแฮร์ริง สารชนิดนี้ คือ 6-(furfuryl-amino) purine ซึ่งมีสูตรโครงสร้างคล้ายอะดีนีน เนื่องจากสารชนิดนี้สามารถกระตุ้นให้เกิดการแบ่งเซลล์โดยร่วมกับออกซิน จึงได้รับชื่อว่าไคเนติน (Kinetin)
ไคเนติน เป็นสารที่ไม่พบตามธรรมชาติในต้นพืช แต่เป็นสารสังเคราะห์ ต่อมาได้มีการค้นพบไซโตไคนินสังเคราะห์อีกหลายชนิด สารสังเคราะห์ที่มีกิจกรรมของไซโตไคนินสูงที่สุดคือ เบนซิลอะดีนีน (Benzyladenine หรือ BA) และเตตระไฮโดรไพรานีลเบนซิลอะดีนีน (tetrahydropyranylbenzyladenine หรือ PBA)
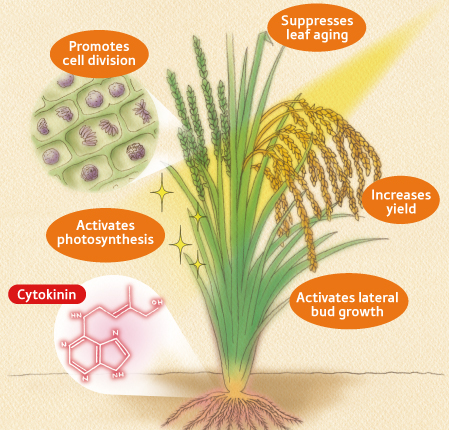
กลไกการทำงานของไซโตไคนิน
ไซโตไคนินมีบทบาทสำคัญคือควบคุมการแบ่งเซลล์ และไซโตไคนินที่เกิดในสภาพธรรมชาตินั้นเป็นอนุพันธ์ของอะดีนีนทั้งสิ้น ดังนั้นงานวิจัยเกี่ยวกับกลไกการทำงานจึงมีแนวโน้มในความสัมพันธ์กับกรดนิวคลีอิค กลไกการทำงานของไซโตไคนินยังไม่เด่นชัดเหมือนกับออกซิน และจิบเบอเรลลิน แต่ไซโตไคนินมีผลให้เกิดการสังเคราะห์ RNA และโปรตีนมากขึ้นในเซลล์พืช ผลการทดลองบางรายงานกล่าวว่า หลังจากให้ไซโตไคนินกับเซลล์พืชแล้วจะเพิ่มปริมาณของ m-RNA, t-RNA และ r-RNA
การศึกษากลไกการทำงานของไซโตไคนิน ในช่วงทศวรรษ 1960 ได้เน้นไปในแง่ที่ว่าไซโตไคนินอาจจะส่งผลของฮอร์โมนผ่าน t-RNA บางชนิด เนื่องจากมีการค้นพบว่ามีกลุ่มไซโตไคนินปรากฏอยู่ร่วมกับ t-RNA หลายชนิด ทั้ง t-RNA ของซีรีน (Serine) และไธโรซีน (Thyrosine) มี อะดีนีนเบสซึ่งมี side chain และมีคุณสมบัติเป็นไซโตไคนินซึ่งมีประสิทธิภาพสูง ยิ่งไปกว่านั้นในกรณี อะดีนีนซึ่งมีคุณสมบัติของไซโตไคนินจะอยู่ถัดจากแอนติโคดอน (Anticodon) ของ t-RNA จึงเป็นที่เชื่อกันว่าการปรากฏของไซโตไคนินบน t-RNA อาจจะจำเป็นต่อการเกาะกันของโคดอน (Codon) และแอนติโคดอนระหว่าง m-RNA และ t-RNA บนไรโบโซม ซึ่งสมมุติฐานที่ว่าไซโตไคนินควบคุมกระบวนการ Translation ผ่านทาง t-RNA จึงได้รับความเชื่อกันมากในขณะนั้น
อย่างไรก็ตามสมมุติฐานนี้ ในเวลาต่อมาได้รับการวิจารณ์อย่างรุนแรง เช่นในการสังเคราะห์ t-RNA ตามปกตินั้น อาจจะเกิดการเปลี่ยนรูปของเบส หลังจากที่มีโพลีนิวคลีโอไทด์ (Polynucleotide) แล้ว ซึ่งหมายความว่า side chain บนตำแหน่งที่ 6 ของอะดีนีนนั้นเกิดขึ้นหลังจากที่อะดีนีนได้อยู่บน t-RNA เรียบร้อยแล้ว ซึ่งเป็นไปไม่ได้ที่ไคเนตินและซีเอตินหรือไซโตไคนินอื่นๆ จะเข้าร่วมกับ t-RNA ในรูปที่เป็นโมเลกุลที่สมบูรณ์ หลักฐานอีกข้อที่ไม่สนับสนุนสมมุติฐานนี้คือ การพบว่า t-RNA ของเมล็ดข้าวโพดซึ่งมีซีส-ซีเอติน (Cis-Zeatin) ในขณะที่ไซโตไคนินที่เกิดในธรรมชาติในเมล็ดเดียวกันเป็นทรานส์-ซีเอติน (trans-Zeatin) ดังนั้นจึงเป็นการยากที่จะเชื่อว่าไซโตไคนินเป็นสารเริ่มต้นของการสังเคราะห์ t-RNA แม้ว่างานทดลองจะยังสับสนและขัดแย้งกัน แต่โดยทั่วไปการรวมของไซโตไคนินเข้าไปใน t-RNA นั้นเกิดในอัตราที่ต่ำมากจนไม่น่าเชื่อสมมุติฐานดังกล่าว
งานวิจัยได้เปลี่ยนแนวและสนับสนุนว่าไซโตไคนินอาจจะทำงานโดยควบคุม กิจกรรมของเอนไซม์โดยตรงมากกว่าที่จะเกี่ยวกับการสังเคราะห์เอนไซม์ ไซโตไคนินมีอิทธิพลต่อเอนไซม์หลายชนิด เช่น ไคเนส (Kinases) ที่ใช้ในกระบวนการหายใจ นอกจากนั้นกิจกรรมของเอนไซม์ที่ใช้ในกระบวนการสังเคราะห์แสงก็เพิ่มขึ้น
ผลของไซโตไคนิน
- กระตุ้นให้เกิดการแบ่งเซลล์และการเปลี่ยนแปลงทางคุณภาพใน tissue culture โดยต้องใช้ร่วมกับ Auxin ในการเลี้ยงเนื้อเยื่อพืชนั้นหากให้ฮอร์โมน ไซโตไคนินมากกว่าออกซิน จะทำให้เนื้อเยื่อนั้นเจริญเป็น ตา ใบ และลำต้น แต่ถ้าหากสัดส่วนของออกซินมากขึ้นกว่าไซโตไคนินจะทำให้เนื้อเยื่อนั้นสร้างรากขึ้นมา การ differentiate ของตาในการเพาะเลี้ยงเนื้อเยื่อจาก Callus จากส่วนของลำต้นนั้น auxin จะระงับ และไซโตไคนินนั้นจะกระตุ้นการเกิด และต้องมีความสมดุลระหว่างไซโตไคนินและออกซินชิ้นเนื้อเยื่อจึงจะสร้างตาได้
- ชะลอกระบวนการเสื่อมสลาย เช่น กรณีของใบที่เจริญเต็มที่แล้วถูกตัดออกจากต้น คลอโรฟิลล์ RNA และโปรตีนจะเริ่มสลายตัวเร็วกว่าใบที่ติดอยู่กับต้น แม้จะมีการให้อาหารกับใบเหล่านี้ก็ตาม ถ้าหากเก็บใบเหล่านี้ไว้ในที่มืดการเสื่อมสลายยิ่งเกิดเร็วขึ้น อย่างไรก็ตามหากใบเหล่านี้เกิดรากขึ้นที่โคนใบหรือก้านใบ จะทำให้การเสื่อมสลายเกิดช้าลง เพราะไซโตไคนินผ่านขึ้นมาจากรากทางท่อน้ำ อย่างไรก็ตามการให้ไซโตไคนินกับใบพืชเหล่านี้จะชะลอการเสื่อมสลายได้เหมือนกับรากเช่นกัน นอกจากนั้นไซโตไคนินยังทำให้มีการเคลื่อนย้ายอาหารจากส่วนอื่นมายังส่วนที่ได้รับไซโตไคนินได้ เช่น กรณีของใบอ่อนซึ่งมีไซโตไคนินมากกว่าใบแก่จะสามารถดึงอาหารจากใบแก่ได้
ในกรณีเชื้อราที่ทำให้เกิดโรคราสนิม ซึ่งทำให้เกิดการตายของเนื้อเยื่อแล้วบริเวณเนื้อเยื่อที่ตายจะเกิดสีเขียวล้อมรอบขึ้นมาซึ่งบริเวณสีเขียวนี้มีแป้งสะสมมากแม้กระทั่งส่วนอื่นๆ ของใบตายไปแล้ว ส่วนสีเขียวอาจจะยังคงอยู่ ลักษณะนี้เรียกว่า Green Island ซึ่งบริเวณนี้จะมี ไซโตไคนินสูง คาดว่าเชื้อราสร้างขึ้นมาเพื่อดึงอาหารมาจากส่วนอื่น - ทำให้ตาข้างแตกออกมาหรือกำจัดลักษณะ Apical Dominanceได้ การเพิ่ม ไซโตไคนินให้กับตาข้างจะทำให้แตกออกมาเป็นใบได้ ทั้งนี้เพราะตาข้างจะดึงอาหารมาจากส่วนอื่นทำให้ตาข้างเจริญได้ เชื้อจุลินทรีย์บางชนิดสามารถสร้างไซโตไคนินกระตุ้นให้พืชเกิดการแตกตาจำนวนมากมีลักษณะผิดปกติ เช่น โรค Fascination นอกจากนั้นยังเร่งการแตกหน่อของพืช เช่น บอน และโกสน
- ทำให้ใบเลี้ยงคลี่ขยายตัว กรณีเมล็ดของพืชใบเลี้ยงคู่งอกในความมืด ใบเลี้ยงจะเหลืองและเล็ก เมื่อได้รับแสงจึงจะขยายตัวขึ้นมา ซึ่งเป็นการควบคุมของไฟโตโครม แต่ถ้าหากให้ไซโตโคนินโดยการตัดใบเลี้ยงมาแช่ในไซโตไคนิน ใบเลี้ยงจะคลี่ขยายได้เช่นกัน ลักษณะดังกล่าวพบกับ แรดิช ผักสลัด และแตงกวา ออกซินและจิบเบอเรลลินจะไม่ให้ผลดังกล่าว
- ทำให้เกิดการสร้างคลอโรพลาสต์มากขึ้น ซึ่งเป็นการเปลี่ยนแปลงทางคุณภาพอย่างหนึ่ง เช่น เมื่อ Callus ได้รับแสงและไซโตไคนิน Callus จะกลายเป็นสีเขียว เพราะพลาสติคเปลี่ยนเป็นคลอโรพลาสต์ได้ โดยการเกิดกรานาจะถูกกระตุ้นโดยไซโตไคนิน
- ทำให้พืชทั้งต้นเจริญเติบโต
- กระตุ้นการงอกของเมล็ดพืชบางชนิด
กรดแอบซิสิค (Abscisic Acid) หรือ ABA
ในการศึกษาถึงการร่วงของใบ การพักตัวของตาและเมล็ดในช่วงปี ค.ศ. 1950-1960 นั้น ชี้ให้เห็นว่าเป็นไปได้ว่ามีสารระงับการเจริญปรากฏอยู่ในต้นพืช โครงสร้างของสารเคมี ดังกล่าวถูกค้นพบในปี 1965 ในผล และใบของฝ้าย สารเคมีดังกล่าวได้รับการตั้งชื่อว่า กรดแอบซิสิค หรือ ABA และพบว่าเป็นสารจำพวกเซสควิเทอร์พีนอยด์
โมเลกุลของ ABA ประกอบด้วย asymmetric carbon atom จึงสามารถแสดงลักษณะของ optical isomerism ได้ อย่างไรก็ตามในสภาพธรรมชาติ ABA จะเกิดเพียงชนิด (+) enantiomorph เท่านั้น ABA ยังแสดงลักษณะ geometric isomerism ได้ด้วย side chain จะเป็น trans รอบๆ คาร์บอนตำแหน่งที่ 5 เสมอ แต่โมเลกุลสามารถเป็นได้ทั้ง cis- หรือ trans รอบๆ คาร์บอนตำแหน่งที่ 2 ABA ส่วนใหญ่ที่พบในพืชจะเป็น (+)-2-cis ABA แม้ว่าจะพบ (+)-2-trans ABA บ้างแต่น้อยมาก ดังนั้นรูป (+)-2-cis ของ ABA จึงมักหมายถึง ABA ทั่ว ๆ ไป
ABA ถูกแยกออกจากพืชหลายชนิดทั้งแองจิโอสเปิร์มส์ จิมโนสเปิร์มส์ เฟินและมอส (Angiosperms, Gymnosperms, Ferns และ Mosses)
กลไกในการทำงานของ ABA
ABA มีกลไกการทำงานคล้ายคลึงกับฮอร์โมนพืชชนิดอื่นๆ คือจะเปลี่ยนระดับและกิจกรรมของเอนไซม์ในรูปเมตาบอลิสม์ของกรดนิวคลีอิค ABA สามารถชะงักการปรากฏของแอลฟาอะมัยเลสในเซลล์อะลีโรนของข้าวบาร์เลย์ได้
ในทำนองเดียวกันกลไกของการทำงานของ ABA คล้ายคลึงกับฮอร์โมนชนิดอื่นๆ คือ เกี่ยวข้องกับการควบคุมเมตาบอลิสม์ของกรดนิวคลีอิค และการสังเคราะห์โปรตีน ซึ่งการควบคุมอาจจะเกิดได้หลายวิธี เช่น ABA กระตุ้นกิจกรรมของเอนไซม์ไรโบนิวคลีเอส (Ribonuclease) หรือ RNase ซึ่งจะทำลาย RNA ทำให้อัตราการสังเคราะห์โปรตีนลดลง แต่อย่างไรก็ตามมีข้อขัดแย้งว่า ABA สามารถลดการสังเคราะห์ RNA ลงได้ภายใน 3 ชั่วโมง หลังจากที่พืชได้รับ ABA แต่จริงๆ แล้วกิจกรรมของ RNase จะเพิ่มขึ้น หลังจากนั้น 8 ชั่วโมง ซึ่งชี้ให้เห็นว่าผลของ ABA ในชั้นต้นไม่ใช่การสังเคราะห์ RNase แต่ยังไม่ทราบแน่ชัดว่าคืออะไร
ในทางตรงกันข้ามกับสภาวะที่ ABA ก่อให้เกิดการลดปริมาณ RNA รวมของเซลล์ มีการพบว่าในเนื้อเยื่ออะลีโรนของข้าวบาร์เลย์นั้นพบว่า การสังเคราะห์แอลฟาอะมัยเลสถูกหยุดชะงัก โดย ABA ไม่มีผลต่อ RNA รวม หรือการสังเคราะห์โปรตีนรวมเลย ผลของ ABA ที่มีต่อแอลฟา อะมัยเลส อาจจะไม่ผ่านมาทางการสังเคราะห์ m-RNA ซึ่งแปลรหัสเพื่อสร้างแอลฟาอะมัยเลสด้วย คาดว่า ABA จะมีผลต่อ regulator RNA ที่ใช้ในกระบวนการ Translation ของแอลฟา อะมัยเลส
ถึงแม้ว่าจะทราบกันดีว่า ABA มีผลต่อการสังเคราะห์โปรตีน อย่างไรก็ตามมีผลของ ABA หลายกรณีที่เกิดขึ้นเร็วเกินกว่าที่จะอธิบายโดยกระบวนการนี้ เช่น การปิดของปากใบ ซึ่งเกิดภายในไม่กี่นาทีที่ได้รับ ABA นอกจากนั้นการยืดตัวของโคลีออพไทล์ที่ได้รับออกซิน จะหยุดชะงักภายใน 2-3 นาที เมื่อได้รับ ABA จึงสรุปได้ว่า ABA มีผลต่อพืชโดยไม่ผ่านเมตาบอลิสม์ของ RNA ได้
ความสัมพันธ์ของ ABA กับการขาดน้ำ
ปริมาณของ ABA ในต้นพืชจะมีความสัมพันธ์โดยตรงกับค่า Water potential ของพืช ปริมาณ ABA ในต้น Ambrosia artemisifolia จะเพิ่มขึ้นทันทีเมื่อใบพืชชนิดนี้มี Water potential เป็น -10 บาร์ ส่วนในข้าวโพดจะอยู่ที่ -12 บาร์ ส่วนในถั่วปริมาณ ABA จะสูงขึ้นเมื่อ Water potential เท่ากับ -7 บาร์
การขาดน้ำทำให้เยื่อหุ้มคลอโรพลาสต์มี Permeability เพิ่มขึ้น ส่งผลให้เกิดการ รั่วไหลของ ABA จากคลอโรพลาสต์ซึ่งเป็นบริเวณที่สะสม ABA ของใบปกติ การลดปริมาณของ ABA ในคลอโรพลาสต์จะกระตุ้นให้มีการสังเคราะห์ ABA เพิ่มขึ้น เมื่อพืชได้รับน้ำเยื่อหุ้มคลอโรพลาสต์จะไม่ยอมให้ ABA รั่วไหลออกมา ทำให้เกิดการหยุดสังเคราะห์ ABA ในคลอโรพลาสต์ ซึ่งเป็น Feedback mechanism
ABA กระตุ้นให้เกิดการปิดของปากใบได้เพราะ ABA จะระงับการผ่านของ K+ เข้าสู่ Guard cell และกระตุ้นให้ Matate รั่วไหลออกจาก Guard cell และนอกจากนั้น ABA ยังระงับการแลกเปลี่ยนประจุ H+ และ K+ ของ Guard cell ทำให้ปริมาณของ K+ และ Malate ใน Guard cell มีน้อย ทำให้ค่า Water potential สูงขึ้น น้ำจึงไหลออกจาก Guard cell ทำให้ปากใบปิดได้
ผลของ ABA ต่อพืช
- ลดการคายน้ำโดยกระตุ้นให้ปากใบปิด ซึ่งพืชตอบสนองได้ภายใน 1-15 นาที หลังจากได้รับ ABA ในพืชที่ขาดน้ำจะมีปริมาณ ABA เพิ่มขึ้นอย่างรวดเร็ว ซึ่งเป็นการลดการคายน้ำของพืช
- กระตุ้นให้เกิดการพักตัวของตา ซึ่งจะเกิดกับพืชเขตอบอุ่น พบว่าเมื่อให้ ABA กับตาที่กำลังเจริญเติบโต จะทำให้ตาชะงักการเจริญเติบโตและเข้าสู่การพักตัวตามปกติ การให้ GA จะลดผลของ ABA ที่ทำให้ตาพักตัวได้
- การร่วงของใบและดอก เช่น ในฝ้าย ผลแก่ที่ร่วงเองจะมี ABA สูงมาก
- เร่งกระบวนการเสื่อมสภาพของใบ
เอทธิลีน (Ethylene)
เป็นฮอร์โมนพืชซึ่งควบคุมการเจริญเติบโตในหลายแง่ เช่น การพัฒนา การเสื่อมสลาย ขึ้นอยู่กับเวลาและสถานที่ ซึ่งเกิดเอทธิลีนขึ้นมา ผลของเอทธิลีนมีทั้งในแง่ที่เป็นประโยชน์หรือเป็นโทษต่อพืช เอทธิลีนเป็นฮอร์โมนที่มีสภาพเป็นก๊าซซึ่งรู้จักกันมานานแล้ว จากการบ่มผลไม้ ในปี 1934 ได้มีการพิสูจน์ให้เห็นว่าเอทธิลีนเป็นก๊าซที่สังเคราะห์ขึ้นโดยพืช และสามารถเร่งกระบวนการสุกได้ ต่อมาพบว่าการก่อกองไฟใกล้ ๆ สวนมะม่วงและสับปะรดจะกระตุ้นให้ออกดอกได้ ซึ่งสารที่ทำให้เกิดการออกดอก คือ เอทธิลีนนั่นเอง เอทธิลีนเป็นฮอร์โมนพืชที่สำคัญในด้านหลังเก็บเกี่ยวด้วย
ต่อมาพบว่า ดอก เมล็ด ใบ และรากพืชสามารถสังเคราะห์เอทธิลีนได้ เช่น ใน เซเลอรี่ พันธุ์ซึ่งต้นขาวเอง (Self blanching) พบว่า เซเลอรี่สามารถสร้างเอทธิลีนมากำจัดสีเขียวที่ก้านได้ นอกจากนั้นในปี 1935 ยังพบว่า การให้ออกซินกับพืชอาจจะกระตุ้นให้พืชสร้างเอทธิลีนได้ ซึ่งเป็นคำอธิบายได้ชัดเจนสำหรับกรณีที่เมื่อให้ออกซินกับพืชแล้วพืช ตอบสนองเหมือนกับได้รับเอทธิลีน ออกซินกับเอทธิลีนนั้นเมื่อให้กับพืชมักจะให้ผลส่งเสริมกัน ส่วนของพืชที่พบเอทธิลีนมากคือ ใบแก่ ผลไม้สุก และเนื้อเยื่อที่อยู่ภายใต้สภาพความเครียด (Stress)
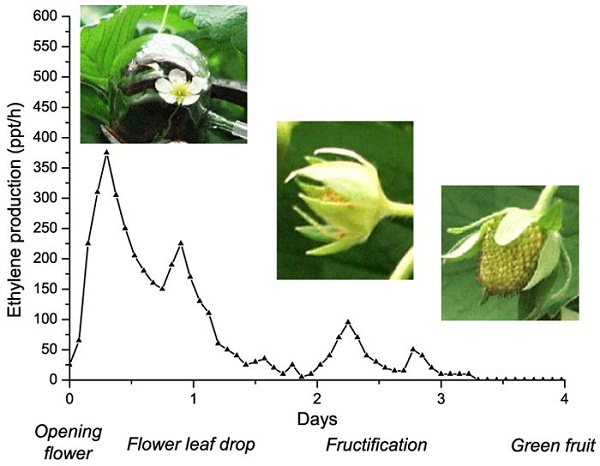
กลไกการทำงานของเอทธิลีน
จากการศึกษาพบว่า เอทธิลีนจะก่อให้เกิดผลต่อการเจริญเติบโตของพืช โดยการรวมกับ receptor site ซึ่งมีโลหะรวมอยู่ในโมเลกุลด้วย และคาดว่าโลหะดังกล่าวคือทองแดงและ analogues ของเอทธิลีนจะสามารถแข่งขันเข้ารวมกับ receptor site และก่อให้เกิดผลคล้ายคลึงกับผลของเอทธิลีน บริเวณที่เอทธิลีนรวมกับ receptor site นั้น คาดว่าอยู่บริเวณเยื่อหุ้มเซลล์หลังจากที่รวมกับ receptor แล้วจะทำให้ receptor สามารถกระตุ้นให้เกิดการตอบสนองขึ้น ซึ่งจะนำไปสู่การตอบสนองขั้นต่อไป
การที่เนื้อเยื่อพืชได้รับเอทธิลีนจะมีการเปลี่ยนแปลงทั้งด้านปริมาณและคุณภาพของเอนไซม์ ดังนั้นจึงเป็นไปได้ว่าเอทธิลีนจะควบคุม RNA ให้สังเคราะห์โปรตีน มีเอนไซม์หลายชนิดมีกิจกรรมเพิ่มขึ้นหลังจากที่ได้รับเอทธิลีนแล้ว เช่น เซลลูเลส (Cellulase) เพอร์ออกซิเดส (Peroxidase) ฟีนีลอะลานีน แอมโมเนีย ไลเอส (Phenylalanine ammonia lyase) และฟอสฟาเตส (Phosphatase) ตัวอย่างของการกระตุ้นให้เกิดเซลลูเลสที่เด่นชัดที่สุดคือการเกิดรอยแยกขึ้นที่ก้านใบกับลำต้น หรือกิ่งในระหว่างการร่วงของใบซึ่งเกิดจากการได้รับเอทธิลีน และการเพิ่มขึ้นของเซลลูเลสเกิดจากการกระตุ้นของเอทธิลีน แต่อย่างไรก็ตามการกระตุ้นการเกิดการร่วงของใบโดยเอทธิลีนนั้นรวดเร็วมากเกินกว่ากลไกของการสังเคราะห์โปรตีน ดังนั้นจึงเป็นไปได้ว่าเอทธิลีนกระตุ้นให้เซลลูเลสที่ปรากฏอยู่ในเซลล์แล้ว ปลดปล่อยออกมายังผนังเซลล์ทำให้เกิดการแยกของเซลล์และใบร่วง
ผลของเอทธิลีนที่เกิดอย่างรวดเร็วนั้น จะไม่สามารถหยุดยั้งด้วย สารระงับการสร้าง RNA หรือโปรตีน เช่น แอคติโนมัยซิน ดี และไซโคเฮกซิไมด์
การที่เอทธิลีนละลายได้ดีมากในไขมัน จึงคาดว่า receptor site ก็อยู่ร่วมกับเยื่อหุ้มเซลล์ ซึ่งลักษณะดังกล่าวจึงทำให้เอทธิลีนมีผลต่อการยอมให้สารเข้าออกจากเยื่อหุ้มเซลล์ด้วย แต่กลไกเหล่านี้ยังสลับซับซ้อนและไม่ทราบเด่นชัดนัก ดังนั้นกลไกการทำงานของเอทธิลีนในระยะสั้นอาจจะเกี่ยวข้องกับเยื่อหุ้มเซลล์ ส่วนในระยะยาวจะเกี่ยวข้องกับการสังเคราะห์โปรตีน

ผลของเอทธิลีนต่อพืช
- กระตุ้นให้ผลไม้สุก ดังนั้นอาจจะเรียกเอทธิลีนว่า Ripening hormone และใช้ในการบ่มผลไม้ในทางการค้า
- กระตุ้นการเปลี่ยนแปลงทางคุณภาพ เช่น กระตุ้นให้เกิด Abcission zone ขึ้น ทำให้ใบและกลีบดอกร่วงได้ กระตุ้นให้เกิดการเปลี่ยนแปลงทางคุณภาพของราก และลำต้น รวมทั้งกระตุ้นการออกดอกของพืช เช่น สับปะรด กระตุ้นให้เกิด Adventitious root
- กระตุ้นให้พืชออกจากการพักตัว เช่น กรณีของมันฝรั่ง
- กระตุ้นให้เกิดดอกตัวเมียมากขึ้นในพืช Dioeciousฮอร์โมนที่กล่าวมาแล้วทั้งหมดนี้ ในปัจจุบันได้นำมาใช้ในทางการเกษตรกันอย่างแพร่หลาย ส่วนสารควบคุมการเจริญเติบโตที่นำมาใช้ประโยชน์ในปัจจุบันก็มีหลายชนิด เช่น Paclobutrazol ซึ่งใช้ควบคุมความสูงของพืช ป้องกันการหักล้มของธัญพืชลดความสูงของไม้ประดับ และยังกระตุ้นให้มะม่วงออกผลนอกฤดู ส่วน Chlormequat สามารถใช้ป้องกันการหักล้มของธัญพืช Maleic hydrazide ซึ่งใช้ระงับการงอกของหัวมันฝรั่งและหอมหัวใหญ่ Ethephon ใช้ในการเร่งการไหลของน้ำยางของยางพาราเป็นต้น
การผลิตน้ำหมักฮอร์โมนพืชปราบศัตรูพืชครอบจักรวาล
เตรียมวัสดุ / ปัจจัยการผลิต
- รวบรวมพืชสมุนไพรที่มีฤทธิ์เป็นยาเช่น ข่า ตระไคร้เทศ ใบ/ยอดสะเดา ใบ/ยอดขี้เหล็ก ต้นกล้วย สาบเสือ บอระเพ็ด พริกเน่า หว่านเกราะเพชร โหระพา เถาตดหมา เป็นต้น
- รวบรวมพืชสมุนไพรที่มีฤทธิ์เป็นยาพิษ เช่น กลอย หางไหล หน่อง เห็ดเบื่อ หว่านอุตพิต ใบยูคาลิปตัส สาบแร้งสาบกา เป็นต้น
- นำพืชทุกชนิดที่เตรียมไว้ (ข้อ 1.1+1.2) สับให้ละเอียดด้วยพร้าเครื่องบดสับคลุกเคล้าให้เข้ากัน กลายเป็นเศษพืช
- กากน้ำตาล
- น้ำ
เตรียมอุปกรณ์
- ถังน้ำหมักพร้อมสายรด ขนาดต่างๆ
- พาย (ขนาดด้ามเสียม) ยาว 1-1.20 เมตร ตามขนาดถังหมัก
- พร้า / มีด หรือเครื่องบดสับ
- กะละมัง
วิธีทำใช้สูตร อัตราส่วน 3 ต่อ 1 ต่อ 10 ดังนี้
- ชั่งพืชที่บดสับเป็นเศษพืช เป็น………….กิโลกรัม เช่น 3 กิโลกรัม เทลงในกะละมัง
- ชั่งกากน้ำตาล เป็น……………ลิตร เช่น 1 ลิตร เทกากน้ำตาลลงผสมเศษพืช (ข้อ 3.1) คนให้ทั่วด้วยไม้พาย สังเกตเข้ากันดีแล้วเศษพืชจะเป็นสีน้ำ จึงเทลงใส่ในถังน้ำหมัก
- เตรียมน้ำ เป็น……………ลิตร เช่น 10 ลิตร เทน้ำลงผสมในถังน้ำหมัก
- ครบ 3 หรือ 7 วัน เปิดฝา ใช้ไม้พายคนให้ทั่วแล้วปิดฝาไว้ตามเดิมสามารถนำไปใช้ได้ เก็บได้นาน 6 เดือน ถึง 1 ปี
วิธีใช้น้ำหมัก
แนะนำการใช้ต่อไร่โดยประมาณ ตามลักษณะอาการที่เป็น น้ำหมัก 20-50 ลิตร ผสมน้ำ 1,000 2,000 ลิตร ฉีดพ่นหรือผสมปุ๋ยคอก 250-500 กิโลกรัม ผสมโดโลไมล์ 50 กิโลกรัม (พอกำหมาด) หรือผสมสารสะเดาผง 10-30 กิโลกรัม หรือบอระเพ็ดสด 10-30 กิโลกรัมหรือฝักคูณ 10-50 กิโลกรัม เป็นต้น หว่านในนาข้าว (ใช้ในเดือนสิงหาคม ร่วมใส่ปุ๋ยครั้งที่ 2) ถ้ามีเพลี้ยต่างๆ หนอนกินใบใช้น้ำมันเครื่องเก่าเทลงน้ำไม่เกิน 1 กระป๋องนม น้ำมันเครื่องจะกระจายทั่วผืนนาอุดตันระบบหายใจของเพลี้ยต่างๆ และ หนอนกินใบ
สรุป
- การนำน้ำหมักไปใช้ผสมปุ๋ยคอกยังไม่มีสูตรตายตัว ใช้ในปริมาณมากก็ไม่เกิดผลเสียแก่พืชเนื่องจากน้ำหมัก คือ อินทรียวัตถุ แนะนำให้เกษตรกรใช้ร่วมกับปุ๋ยคอกหรือปุ๋ยขี้ไก่ขี้หมูเพราะน้ำหมักฯ มีธาตุอาหารน้อยมากต้อง ต้นข้าวจะแข็งแรงต้านทานโรคได้
- น้ำหมักฯ ไม่ใช่ปุ๋ยแต่มีฤทธิ์ทางยา ขับไล่แมลงหรือป้องกันการลอกครอบหรือเบื่อเมากัดกินพืชไม่ได้ ตายไปเอง
ป้ายคำ : จุลินทรีย์, ปุ๋ยหมักชีวภาพ
เรื่องที่เกี่ยวข้องกับหมวด ภูมิปัญญา
-
 ปกติการทำ ซั้ง ทำได้หลากหลายวิธีการ จะทำเป็นประการังเทียมก็ได้ หรือทำจากวัสดุท้องถิ่นที่สามารถย่อยสลายในน้ำได้ เช้นเถาวัลย์ ทางมะพร้าว ฯลฯ ซั้งเชือก คือหญ้าเทียมในทะเล ...
ปกติการทำ ซั้ง ทำได้หลากหลายวิธีการ จะทำเป็นประการังเทียมก็ได้ หรือทำจากวัสดุท้องถิ่นที่สามารถย่อยสลายในน้ำได้ เช้นเถาวัลย์ ทางมะพร้าว ฯลฯ ซั้งเชือก คือหญ้าเทียมในทะเล ... -
 พลังที่สำคัญที่สุดในการแก้ไขปัญหาความยากจนคือพลังของคนในครัวเรือนผู้ซึ่งเป็นเจ้าของปัญหาเอง โดยหัวใจสำคัญคือครัวเรือนต้องรู้จักตนเอง รู้ที่จะปรับความคิดในการพิชิตความจน ...
พลังที่สำคัญที่สุดในการแก้ไขปัญหาความยากจนคือพลังของคนในครัวเรือนผู้ซึ่งเป็นเจ้าของปัญหาเอง โดยหัวใจสำคัญคือครัวเรือนต้องรู้จักตนเอง รู้ที่จะปรับความคิดในการพิชิตความจน ... -
 สระพวง เป็นชุดสระน้ำ ที่มีสระแม่ สระลูก สระหลาน ต่างขนาดกัน ไล่เรียงกันไปเป็นขั้นๆ แต่เชื่อมต่อกัน โดยอาศัยหลักการน้ำไหลจากที่สูงลงที่ต่ำ ...
สระพวง เป็นชุดสระน้ำ ที่มีสระแม่ สระลูก สระหลาน ต่างขนาดกัน ไล่เรียงกันไปเป็นขั้นๆ แต่เชื่อมต่อกัน โดยอาศัยหลักการน้ำไหลจากที่สูงลงที่ต่ำ ...